

Mating in the Shadows of the Welfare State
Evolutionary Game Theory and the Rise of Alternative Reproductive Tactics


Across the animal kingdom, natural selection has sculpted a remarkable array of reproductive strategies, but humans are unique in that we consciously design institutions that shape incentives and constrain behavior. Among institutions, the modern welfare state highlights this dynamic. As resources are redistributed for paternalistic purposes, the State increasingly supplants functions once anchored in family and kinship. By altering costs and benefits, these interventions create conditions for alternative reproductive strategies to emerge.
The patterns of reproductive behavior are illuminated by Robert Trivers’s Parental Investment Theory, which demonstrates how sexes differ in their approaches to the trade-off between parental care and mating efforts. Applicable across species, the theory predicts that the sex that invests more in parenting (e.g., gestation, lactation, food provisions, or training) will be more selective when choosing a mate, while the opposite sex competes for intrasexual mating opportunities. Selection pressures arising from this competition favor conditional responses, which over time, and across variations, can stabilize as Alternative Reproductive Tactics (ARTs) within a population.
Against this evolutionary backdrop, Homo sapiens harness their cognitive capacities to an unmatched degree in reproductive strategies and beyond, employing high-level abstractions and acting purposefully through reason to achieve multi-layered ends. Other intelligent species exhibit elements of culture and technology, but nothing rivals humanity’s achievements. In an effort to impose order, mankind has devised systems in which authority spans from centralized to decentralized, with forms of governance ranging from ruthless tyrants to libertarian societies rooted in natural rights. Along this continuum, modern welfare states justify redistributive policies under the guise of benevolence—and by doing so, subvert incentives that hold families together.
As Hans-Hermann Hoppe observed in Democracy: The God That Failed:
Every form of government welfare—the compulsory wealth or income transfer from “haves” to “havenots” lowers the value of a person’s membership in an extended family-household system as a social system of mutual cooperation and help and assistance. Marriage loses value.
In other words, by systematically altering the payoff matrix of reproduction, welfare states not only redistribute resources; they predictably distort incentives, weaken pair-bonding, and favor reproductive tactics that externalize parental costs, mirroring parasitism in the biological sense. These effects are not pathologies of character, but consequences of institutional design. To analyze how relative incentives are reshaped, we turn to game theory.
Game Theory
In 1944, John von Neumann and Oskar Morgenstern formalized classical game theory in Theory of Games and Economic Behavior. A classical game involves two or more rational actors and can be competitive or cooperative. The strategic interactions among players are often represented in a payoff matrix, where payoffs depend on the strategies each player employs. The prisoner’s dilemma is perhaps game theory’s most famous application, but it has been applied across all walks of life, from couples negotiating conflicting preferences over where to go on a date, to residential segregation, to decisions throughout sports.
A notable extension of this approach is evolutionary game theory, which applies to biological systems and provides a framework for analyzing frequency-dependent evolution of phenotypes. Rather than modeling rational actions, as in classical game theory, it examines how strategies—genetically inherited, conditional, or learned—evolve and perform over time. Payoff matrices capture an organism’s survival and reproductive strategies, molded by natural selection, and measured as fitness. In 1973, John Maynard Smith and George Price introduced the Hawk-Dove game to illustrate the logic of conflict over resources in animals.
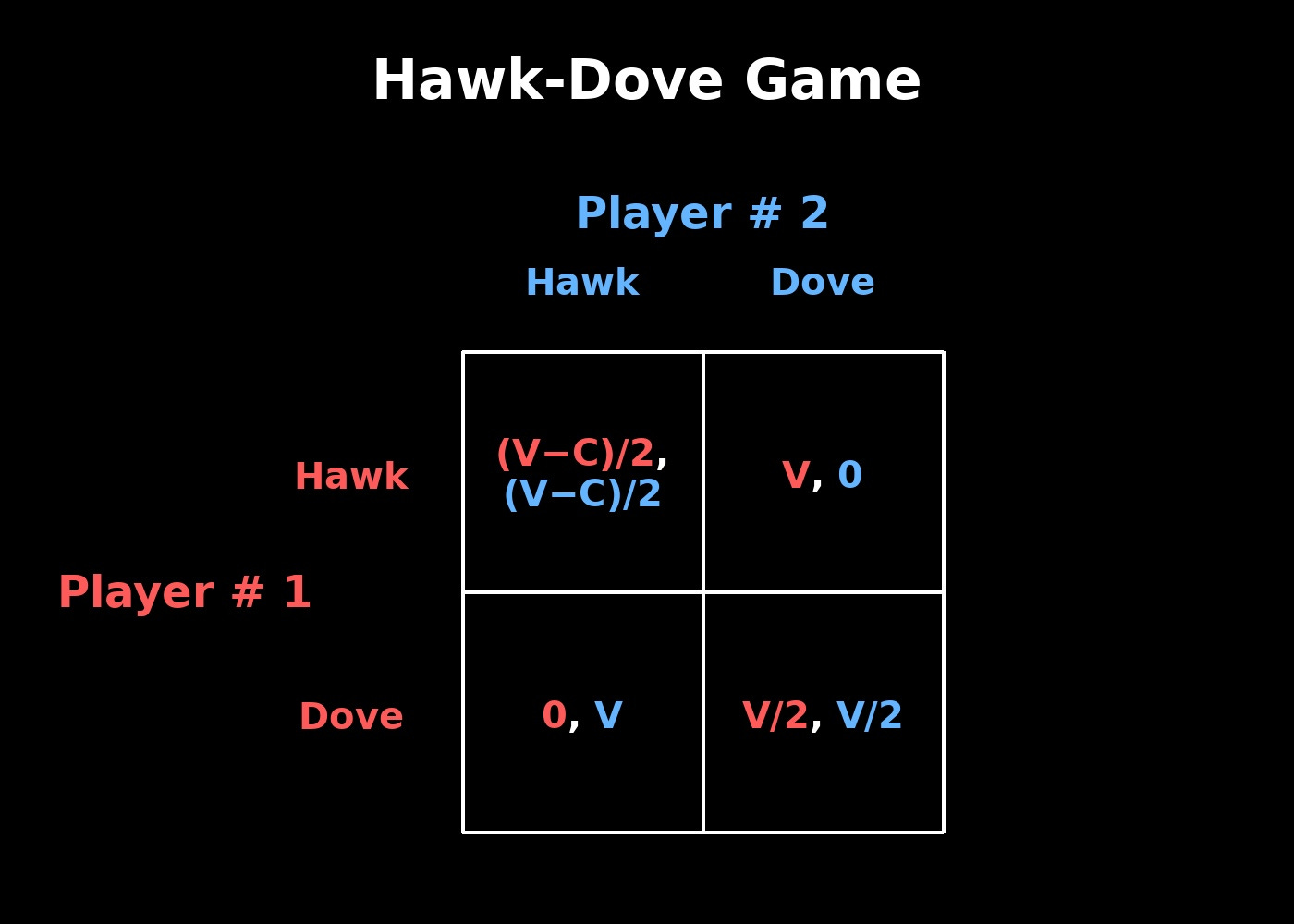
Figure 1. Payoffs are listed as Player #1, Player #2. Colors correspond to each player.
Organisms of the same species engage as players in this game. In each interaction, a player can either escalate aggression (Hawk) or retreat (Dove), with the optimal choice depending on their opponent’s strategy. When one player plays Dove and the other Hawk, the Dove yields while the Hawk claims the resource, producing payoffs of (V,0) or (0,V). If both players play Hawk, conflict ensues, reducing payoffs to (V-C)/2 for each due to the risk of injury. If both players play Dove, the resource is shared equally, (V/2, V/2).
The payoff matrix in the Hawk-Dove game is determined by the resource’s value and the cost of fighting. When V < C, no pure dominant strategy will exist, and the population stabilizes as a mixture of Hawks and Doves. As the resource’s value rises, so does the frequency of Hawks; when the cost of fighting rises, Doves become more common.
Successful strategies, whether genetic, conditional, or learned, replicate and spread according to their relative fitness. Strategies emerge without conscious design, driven by the relentless logic of survival and reproduction. In this sense, much like competing firms in a free market, advantageous strategies proliferate, while disadvantageous ones fade away. Over generations, these pressures harden into enduring behavioral patterns, paving the way for Alternative Reproductive Tactics.
Alternative Reproductive Tactics
Except in asexual species, males and females are the vehicles that pass genes from one generation to the next. In the creation of an embryo, each sex holds a vital resource sought by the other: sperm requires eggs, and eggs require sperm. In the natural world, a dizzying variety of strategies has evolved in the battle to secure mates. When selection pressures produce a discontinuous distribution of behavioral, physiological, or morphological traits—and there is fitness to be gained by pursuing divergent reproductive strategies—an ART may emerge. Given the natural limitations of energy, time, and resources, each tactic carries its own biological costs and benefits. Echoing Thomas Sowell’s insight, evolutionary systems offer no costless solutions—only trade-offs.
Southern elephant seals call the Antarctic home. Males come in two varieties that differ in morphology and reproductive strategy. Bull elephant seals tip the scales at nearly 9,000 lbs, and during mating season, a “beach master” can command a harem of up to 150 females. These beasts roar, bite, and slam their bodies into any and all competitors, exemplifying a high-investment reproductive tactic. Meanwhile, smaller “sneaky” males lurk at the edges of the territory, waiting for the beach master to be distracted so they can slip under his nose and copulate.
Male side-blotched lizards in the southwestern deserts of the US come in three flavors. Each sports its own color and strategy for mating. As with the elephant seal, the orange variety is territorial and polygamous, whereas yellow ones are non-territorial and employ a sneaky strategy. Blue lizards are mostly monogamous, dedicating their resources to mate-guarding while patrolling smaller territories. Akin to rock, paper, scissors, color dominance shifts over time as success is based on the frequencies of competing colors within the population. When orange lizards are common, blue ones struggle to secure mates, and sneaky yellows thrive. As blue lizards become more numerous, sneaky yellows falter, and the cycle repeats.
Throughout North American freshwater lakes, ponds, and streams, bluegill sunfish build colonial circular nests. Larger, nest-guarding males defend the eggs, resulting in high fertilization success. Outside the nests, smaller sneaky males take advantage of lapses in defense during spawning. When the time is right, they dart in, releasing their sperm in a bold bid to fertilize the eggs. Parasitic tactics externalize costs, allowing sneaky males to exploit the energetic and risk-related investments of more committed competitors, though at the cost of lower fertilization success per attempt.
Within a species, high-investment and parasitic strategies fluctuate dynamically over time, often stabilizing due to frequency-dependent selection. Parasitic tactics cannot dominate indefinitely; their success depends on the continued presence of high-investment competitors. When parasitic strategies spread too widely, selection can lock populations into individually adaptive yet collectively maladaptive states, leaving the system destabilized, heightening the risk of population collapse. In non-human species, these dynamics unfold through biological selection alone; in humans, they are further shaped by cognition, culture, and the institutions they construct.
Homo sapiens
Unlike their animal brethren, humans act purposefully through high-level reasoning, using available means to achieve desired ends. The perceived value of potential mates is inherently subjective, shaped by personal histories, interactions, cultures, and societal structures. Individual preferences can be inferred from observable human action. Yet beneath the surface, humanity’s biological underpinnings remain present, as sexual desires can cloud reason. Consequently, in humans, payoff matrices do not represent biological fitness alone, but a hybrid of evolutionary pressures and reasoned decision-making, where strategies respond dynamically to environmental and social conditions.
In terms of parental investment theory, a profound asymmetry exists between the sexes. Women invest at least nine months in pregnancy, and often bear the bulk of post-birth care, while a man’s minimal contribution is measured in minutes and milliliters of semen. As a result, women tend to be more selective when choosing mates, whereas men face competition with other men for reproductive opportunities. These differences in investment influence the conditions under which ARTs emerge in both sexes, though cultural and social factors can modulate their expression.
In contemporary societies, men employ a variety of reproductive strategies. Through a game-theoretic lens, mates function as a proximate resource, instrumental to the goal of offspring production. Monogamous pairings and marriages then act as forms of mate-guarding. Aggression, building on the Hawk-Dove game, can serve as a reproductive tactic. However, aside from its moral reprehensibility, forced copulation—which is descriptive, not justificatory—yields low long-term success due to the significant costs it entails, including injuries, retaliation, ostracism, or incarceration. Alternatively, enabled by technology, a man seeking to maximize his offspring could adopt the sperm bank approach. Other ARTs may emerge as a result of institutional interference.
The rise of institutional strategies is an adaptive response to the incentive structures imposed by their environment. Welfare programs vary widely in eligibility and benefits, and their effects are shaped by prevailing cultural stigmas. For the purposes of this discussion, the safety net refers to assistance—cash or in-kind—provided to impoverished parents. Operating broadly, these institutions systematically replace or supplement support traditionally provided by kinship and pair-bonding, acting as a form of social insurance. By externalizing a portion of parental investment, the safety net reduces penalties associated with non-investment, creating conditions that allow parasitism to take hold as a viable reproductive strategy.
From an evolutionary perspective, a man’s reproductive success benefits from philandering. In a welfare state, however, men who once needed to invest heavily in resource acquisition to attract mates can now redirect their effort toward spreading their genes, as the redistribution of resources reduces the relative cost of abandonment within the payoff matrix. This, in turn, further favors a more polygamous reproductive strategy. Males adopting this parasitic strategy not only gain access to sexual variety but also have the option to adopt a mixed strategy, in which time and resources are selectively provided to improve the well-being of progeny.
From the female perspective, quality of mate is prioritized over quantity. In the absence of a safety net, extreme caution must be exercised when selecting a mate, with emphasis on the man’s ability to assist with child-rearing. The introduction of a welfare system can warp the female payoff matrix, altering the incentives that shape mate choice. Consequently, finding a good, stable provider to support offspring becomes less critical for welfare recipients. As predicted by Gary Becker’s economic analysis of the family, this phenomenon has been termed the “independence effect.” If a woman has her own resources, she may adjust her mate preferences or opt to raise children alone.
The Seattle-Denver Income Maintenance Experiments (SIME-DIME) of the 1970s provide a vivid illustration of the independence effect. Families with at least one dependent below a specified income threshold received guaranteed cash transfers for up to five years. Analyses of the program found significantly higher divorce rates in the experimental group than the control group—approximately 36% higher for black couples and 40% for white couples. Relieved of financial dependency on their spouses, participants adjusted their reproductive strategies.
Since the implementation of the Great Society programs, welfare has dramatically altered family structures. The proportion of births to unmarried women has risen sharply, from 5.3% in 1960 to 40.0% in 2023. At the same time, the share of children living in two-parent households in the United States has declined—from 87.7% in 1960 to 71.1% in 2023. These patterns are disproportionately concentrated among poorer and working-class families, who are also the primary recipients of welfare. While the expansion of the welfare state is only one of many influences and is not a monocausal explanation for these trends, the correlation between state interventions and a reduced reliance on stable pair-bonds is striking.
When resources are subsidized by the State, provisioning becomes less salient in mate selection, and relative rankings of potential partners may shift. As a result, welfare programs distort how subjective values are expressed through demonstrated preference in reproduction. The introduction of institutional ARTs disrupts equilibrium patterns that have historically favored high-investment pair-bonding, facilitating the proliferation of parasitic strategies that undermine traditional families.
Conclusion
From an evolutionary perspective, the welfare state reshapes human incentives. By reducing the personal costs of child-rearing and parental investment, it enables alternative reproductive strategies that would otherwise remain marginal. Over time, interventions that redistribute resources warp the payoff matrix, fostering dependency on the State while weakening traditional pair-bonding.
These are not the consequences of moral failings, but follow the logic of incentives. When the costs of reproduction are systematically externalized, parasitic strategies gain a relative advantage on the fitness landscape. As long as this payoff advantage persists, low-investment strategies will increase in frequency relative to high-investment strategies, creating conditions that can destabilize the population over generations.
Historically, families and local communities bore primary responsibility for dependents. In such environments, the costs and benefits associated with child-rearing were largely internalized within households and kin networks, keeping reproductive incentives aligned with parental investment.
The welfare state cannot eliminate reproductive trade-offs; it can only select which strategies prevail.
Likes, comments, and shares are always appreciated, as they help new readers discover my work.
All articles are free. If you find value in my writing, consider supporting it with a paid subscription or a crypto donation.
XMR: 8BFN1EyJCsqXUg2NxeYvjDJkajCcTjeMx1TCyF3ssyvHM3JhCfCxkLw6Bheo7zCLiZeyXTHHcQTJD5civHEgzwocH13FdSk

BTC: bc1qj9aqksvruyncnvf7pedafrmal6ffk9ujwauqgl

ETH: 0xE404A29Cb1129652dEDeaA6642B668B763d44FD4

Interesting article. Good breakdown of a complicated topic.